Принципиальное макроструктурное различие двух типов нуклеиновых кислот состоит в том, что ДНК - это единая двойная спираль, то есть макромолекула из двух комплементарно связанных полимерных тяжей, спирально закрученных вокруг общей оси (см. [ 1 , 3 ]), а РНК - однотяжевой полимер. В то же время взаимодействия боковых групп - азотистых оснований - друг с другом, а также с фосфатами и гидроксилами сахаро-фосфатного остова приводят к тому, что однотяжевой полимер РНК сворачивается на себя и скручивается в компактную структуру [ 4 ], подобно сворачиванию полипептидной цепи белка в компактную глобулу. Таким способом уникальные нуклеотидные последовательности РНК могут формировать уникальные пространственные структуры.
Впервые специфическая пространственная структура РНК была продемонстрирована при расшифровке атомной структуры одной из тРНК в 1974 г. [ 5 , 6 ] (рис. 3). Сворачивание полимерной цепи тРНК, состоящей из 76 нуклеотидных мономеров, приводит к формированию очень компактного глобулярного ядра, из которого под прямым углом торчат два выступа. Они представляют собой короткие двойные спирали по типу ДНК, но организованные за счет взаимодействия участков одной и той же цепи РНК. Один из выступов является акцептором аминокислоты и участвует в синтезе полипептидной цепи белка на рибосоме, а другой предназначен для комплементарного взаимодействия с кодирующим триплетом (кодоном) мРНК в той же рибосоме. Только такая структура способна специфически взаимодействовать с белком-ферментом, навешивающим аминокислоту на тРНК, и с рибосомой в процессе трансляции, то есть специфически "узнаваться" ими.

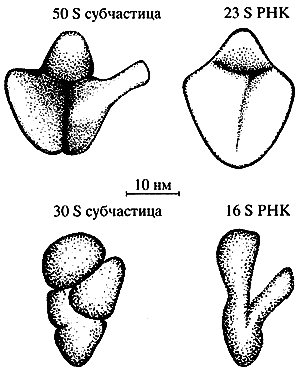
Изучение изолированных рибосомных РНК дало следующий разительный пример формирования компактных специфических структур из еще более длинных линейных полимеров этого типа. Рибосома состоит из двух неравных частей - большой и малой рибосомных субчастиц (субъединиц). Каждая субчастица построена из одной высокополимерной РНК и целого ряда разнообразных рибосомных белков. Длина цепей рибосомных РНК весьма значительна: так, РНК малой субчастицы бактериальной рибосомы содержит более 1500 нуклеотидов, а РНК большой субчастицы - около 3000 нуклеотидов. У млекопитающих, включая человека, эти РНК еще больше - около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно.
Было показано, что изолированные рибосомные РНК, отделенные от их белковых партнеров и полученные в чистом виде, сами способны спонтанно сворачиваться в компактные структуры, по своим размерам и форме похожие на рибосомные субчастицы [7 ]. Форма большой и малой субчастиц разная, и соответственно различается форма большой и малой рибосомных РНК (рис. 4). Таким образом, линейные цепи рибосомной РНК самоорганизуются в специфические пространственные структуры, определяющие размеры, форму и, по-видимому, внутреннее устройство рибосомных субчастиц, а следовательно, и всей рибосомы.
Минорные РНК.
По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" [ 8 ].
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Рибозимы.
Вся активная жизнь построена на обмене веществ - метаболизме, и все биохимические реакции метаболизма происходят с надлежащими для обеспечения жизни скоростями только благодаря высокоэффективным специфическим катализаторам, созданным эволюцией. На протяжении многих десятилетий биохимики были уверены, что биологический катализ всегда и всюду осуществляется белками, называемыми ферментами, или энзимами. И вот в 1982-1983 гг. было показано, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью [ 9 , 10 ]. Такие РНК-катализаторы были названы рибозимами. Представлению об исключительности белков в катализе биохимических реакций пришел конец.
В настоящее время рибосому тоже принято рассматривать как рибозим. Действительно, все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками. Идентифицирован каталитический участок большой рибосомной РНК, ответственный за катализ реакции транспептидации, посредством которой осуществляется наращивание полипептидной цепи белка в процессе трансляции.
Вирусные РНК.
Кроме нуклеиновых кислот (ДНК и РНК), организующих и обслуживающих жизнь клеточных организмов, в природе существуют паразитические молекулы ДНК и РНК. Одетые в защитную белковую оболочку, они называются вирусами. Соответственно, вирусы подразделяются на ДНК-содержащие и РНК-содержащие. Собственно в самих вирусных частицах никакой жизни нет - это просто способ упаковки, консервации и распространения внеклеточного генетического вещества. При попадании в живую клетку вирусная белковая оболочка сбрасывается, а генетическое вещество - нуклеиновая кислота - начинает функционировать как паразит, направляя жизнь клетки на синтез белков, ею кодируемых, и на репликацию самой себя. Так называемое "размножение" вирусов в клетке есть производство многочисленных копий вирусной ДНК или РНК путем репликации, с последующим их "одеванием" в оболочку из синтезированных клеткой вирусных белков.